Introduction
The first Moss rose probably originated at Carcassonne, France, about the year 1696, as a sport of the then much cultivated Cabbage rose (Hurst and Breeze, 1922)
The Moss rose (R. centifolia muscosa), is almost identical with the Cabbage rose ( R . centifolia L.), except that its stems, petioles, pedicels, peduncles, calyx tubes and sepals are copiously covered with glandular mossy protuberances, which give off a resinous or balsamic odour when bruised (after Hurst and Breeze 1922)
Three Moss sports are reported, and all later Moss roses have been derived from them, either directly by continued sporting, or indirectly by hybridyzing. Just like their ancestors, Moss roses are tetraploids, 2n = 4x = 28 (Hurst 1941a).
Probably owing to their very tall bushes and non-recurrent flowering habit, Moss roses have been replaced by the more profusely blooming Hybrid Tea-,Polyantha-, and Floribunda classes of roses.
With the aim to make the attractiveness of the moss habit available in recurrent flowering plants, 1st and 2nd generation crosses between Hybrid Tea and Moss cultivars were made at IVT between 1976 and 1979. The present study reports on the inheritance of the recurrent flowering and moss characters in these crosses
Materials and Methods
From the collection of old rose cultivars at the Wageningen Arboretum, 5 Moss cultivars were selected which, according to SHEPHERD (1954) and Meikle (1980) possibly flowered more than once: 'Marie de Blois' (1852), 'Herman Kegel' (1848), 'William Lobb' (1855), 'Jeanne de Montfort' (1851) and 'Salet' (1854). Pollen from these cultivars was collected in 1976 and used to pollinate the recurrent flowering tetraploid (2n = 4x = 28) Hybrid Tea cultivars: 'Ann Cocker', 'Lara', 'Muria', 'Polynesian Sunset' and 'Predila'.
Pollen of 'Jeanne de Montfort' failed to induce fruit set, but with the other Mosses fruit set of the Hybrid Teas was normal. F1 seedlings did not flower until the 2nd year after germination (1978) and then but once, indicating that they were non-recurrent. Moss, which can only be assessed at flower, varied considerably within F1 populations. Selection was made for the heaviest moss available. In F1 populations with 'Marie de Blois', the moss character was not expressed sufficiently, but 3 F1 seedlings from 'Salet', 7 from 'Herman Kegel', and 11 from 'William Lobb' with pronounced moss characters were chosen to raise a F2 by self-pollination.
Spring 1979, selection was made in F2 populations for recurrent flowering (moss) seedlings; the non-recurrent seedlings, in which moss could not be assessed until 1980, were discarded
Seeds obtained in these crosses were always stratified in humid soil at + 0,5° C for 4 months
Results and Conclusions
Table 1 summarizes the number of non-recurrent and recurrent, and non-moss and moss seedlings per Moss rose pollinator. All F1 populations were uniformly non-recurrent flowering, but they segregated into non-moss and moss plants. F2 populations segregated into non-recurrent and recurrent flowering seedlings, the latter ones into non-moss and moss plants.
Recurrent Flowering
Semeniuk (1971a, 1971b) and DE VRIES and DUBOIS (1978) demonstrated that recurrent flowering in roses is a monogenic character, the non-recurrent allele R being dominant over the recurrent one r. Consequently, in Hybrid Tea cultivars recurrent flowering is controlled by rrrr.
Because in F1 populations no recurrent seedlings occurred, and in F2 populations the ratios non-recurrent: recurrent seedlings differed very litte (Table 1), it may be assumed that, at least in 'Saleť, 'Herman Kegel', and 'William Lobb', recurrent flowering is controlled by the same make-up, i. e. either RRRR or RRRr.
In the former case F1's would be uniformly RRrr, segregating in the F2 into 35 R: 1 r, which does not agree at all with the ratios obtained (Table 1). In the latter case, F1's would consist of 50% RRrr and 50% Rrrr, segregating in the F2 into 35 R: 1 r and 3 R: 1 r respectively or, on average 6.2 R: 1 r
As probabilities obtained in Chi-square analysis indicate a reasonable to very good fit for this ratio (Table 1), it is concluded that recurrent flowering in the Moss cultivars 'Saleť, 'Herman Kegel', 'William Lobb', and probably in 'Marie de Blois' as well, is controlled by RRRr.
Moss
Hurst and Breeze (1922) and Moore (1978) suggested that the moss character in old Moss cultivars might be controlled by a single dominant gene, named M. According to this concept, in Hybrid Tea cultivars moss is controlled by mmmm, in Moss cultivars by M(...)
In the F1 populations described here the 1:1 ratio of non-moss : moss seedlings indicated a probable Mmmm control of moss in the Moss cultivars (Table 1). Chi-square analysis yielded probabilities that show a reasonable to good fit for this hypothesis
Upon selfing, Mmmm F1 plants yielded a F2 of 25% MMmm (moss), 50% Mmmm (moss) and 25% mmmm (non-moss), so a non moss: moss ratio of 1:3. Probabilities obtained in Kaphi-square analysis, indicate a good to very good fit for this ratio (Table 1).
It is thus concluded, that in the Moss rose cultivars 'Saleť, 'Herman Kegel', 'William Lobb* and most probably in 'Marie de Blois' also, moss is controlled by Mmmm. Table 1 also shows that moss and recurrent flowering are unlinked characters
Although the segregation non-moss: moss was quite clear cut, the moss character varied considerably in intensity and could be classed from 1 -9 as follows: 1 = no moss, 2 = very little moss, 9 = very heavy moss (see fig. 1). The distribution of the 92 'William Lobb' F2 seedlings over these classes of moss is illustrated in Fig. 2.
The distribution pattern strongly suggests the presence of modifiers with an apparent additive effect.

Figure 1:(above). Classes of moss on the sepals of seedlings in Hybrid Tea x Moss F2-population
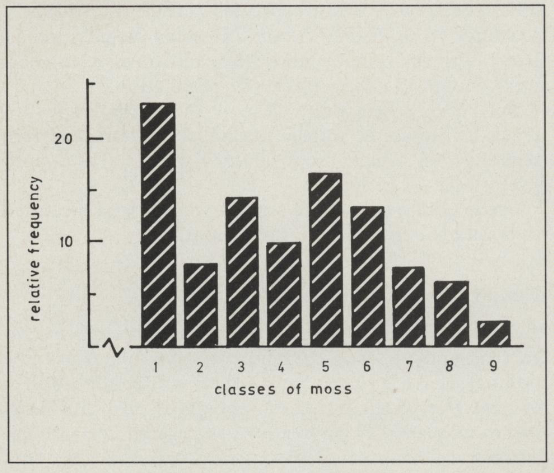
Figure 2:(above). Classes of moss in 11 Hybrid Tea x 'William Lobb' F2-populations. (1 = no moss, 2 = very little moss, 8 = heavy moss, 9 = very heavy moss; n = 92).
Discussion
Present results demonstrated that moss is a single gene, dominant character. This means that the 'moss-sport' must have occurred as a dominant mutation, probably on a single gene basis as well. In rose, where single gene characters are rare anyway (DE Vries and DUBOIS 1978), no other examples of single gene mutation are known.
According to BROCK (1977), even in induced mutations, single gene visible mutants occur at very low rates, the dominant type being about 200 times rarer than the recessive one.
The Moss cultivars used in this experiment have a status as 'more-than-once-flowering' (SHEPHERD 1954, Meikle 1980), although the former author mentions that recurrent flowering in Moss roses is rarely exhibited. Also in the Wageningen Arboretum they flower in spring only.
In view of the above results, which show the presence of only one gene for recurrent flowering in these cultivars, this is not unexpected.
However, genes for recurrent flowering do not originally occur in R. centifolia L. nor in any other European rose. They were only available after the introduction of the four so called 'Stud-Chinas', in England at the beginning of the last century (HURST 1941b, WYLIE 1954). It is concluded therefore, that the one gene for recurrent flowering in the tetraploid Moss cultivars, which all originated around the year 1850, has been transmitted to them from the China source (via a triploid stage) in at least 2 generations of breeding.
It was demonstrated that moss and recurrent flowering may be combined in 2 generations of breeding. The recurrent flowering Moss selections obtained, open the way for future breeding. Most of these selections are easy to breed with, which is convenient because many characters still need improvement
Both in crosses among themselves and with cultivars, recurrent flowering will be preserved. However, as selections are not yet homozygous for moss, populations will always segregate, the ratios depending on the progenitor being simplexes or duplexes, and whether inbreeding or outbreeding is carried out. When rapid improvement is aimed at, outbreeding with desirable cultivars should be preferred rather than inbreeding with Mosses
Summary
In F1- and F2-crosses between 5 tetraploid (2n = 4x = 28), recurrent flowering non-moss Hybrid Tea rose cultivars and tetraploid, non-recurrent flowering Moss rose cultivars, inheritance of the recurrent flowering (r) and moss (M) characters were studied.
Like in the roses studied previously, recurrent flowering is controlled by a single recessive gene r. Moss is controlled by a single dominant gene M; in the ancient Cabbage roses this gene arose as a rare, dominant mutation. The Moss cultivars 'Marie de Blois', 'Herman Kegel','William Lobb', and 'Salet' are triplexes for R and simplexes for M. Breeding with Moss roses is discussed.